Fotogalerie
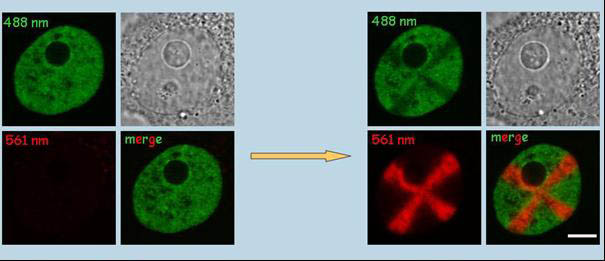 HepG2 buňky exprimující rekombinantní protein histon H4-Dendra2.
HepG2 buňky exprimující rekombinantní protein histon H4-Dendra2.
Neaktivovaná Dendra2 (vlevo) má zelenou fluorescenci (488 nm).
Použitím laseru (405 nm) byla Dendra 2 fotokonvertována v oblasti jádra ve formě kříže (vpravo).
Fotokonverze měla za následek fluorescenci v červené barvě (561 nm), zatímco zelená fluorescence byla v
místech fotokonverze zeslabená. Měřítko, 1µm. (Cvačková et al., J Struct Biol. 2009;165:107-17)
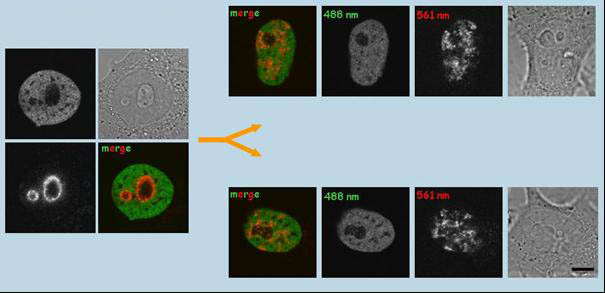 Fotokonverze Dendry 2 asociované s histonem H4 v místech výskytu chromatinu asociovaného
s jadérkem v HepG2H4-Dendra2 buňkách a sledování označené domény v dceřiných buňkách.
Fotokonverze Dendry 2 asociované s histonem H4 v místech výskytu chromatinu asociovaného
s jadérkem v HepG2H4-Dendra2 buňkách a sledování označené domény v dceřiných buňkách.
V experimentu byl histon H4 vybrán proto, že setrvává na stejném segmentu DNA mnoho hodin. Pomocí fluorescenčně
označených částí chromatinu (vlevo) v S/časné G2 fáze bylo na živých buňkách sledováno, do jaké míry je zachována
pozice označené domény po buněčném dělení (vpravo). V dceřiných buňkách (14 hodin po fotokonverzi v mateřské buňce),
je rozmístění vybraných oblastí výrazně jiné, i když ne zcela náhodné. Měřítko, 1µm. (Cvačková et al.,
J Struct Biol. 2009;165:107-17)
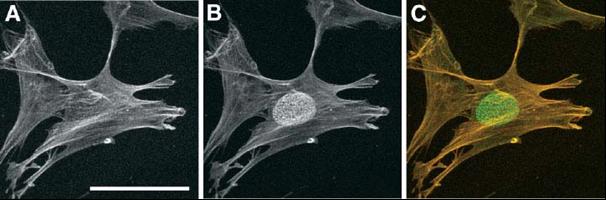 Na chladu závislá detekce nukleárního epitopu epiC.
Na chladu závislá detekce nukleárního epitopu epiC.
Metanolem fixované lidské fibroblastové buňky (LEP) byly inkubovány s myší anti-aktinovou monoklonální
protilátkou 2 hod při RT a následně s Cy3-konjugovanými kozími anti-myšími protilátkami (červený kanál, A)
40 min při RT. Po několikerém oplachu byly buňky inkubovány opět s anti-aktinovou protilátkou při 4°C přes
noc a následně s Alexa Fluor488-kozími anti-myšími protilátkami 60 min (zelený kanál, B). Překryv obou kanálů
je ukázán v (C). Prokázali jsme, že jaderný epitop epiC odpovídá dvojité posttranslační
modifikaci histonu H4. Měřítko, 50µm. (Fidlerová et al., J Cell Biochem. 2005; 94:899-916 a Fidlerová et al., J Struct Biol.
2009 Apr 5. [Epub ahead of print])
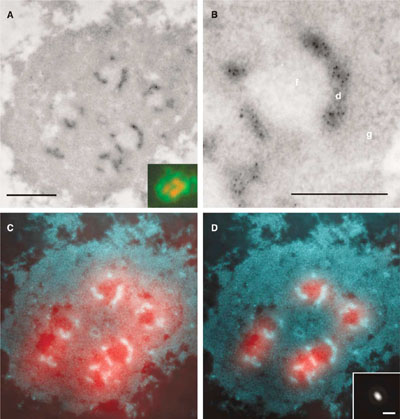 Lokalizace transkripce ribozomálních genů.
Lokalizace transkripce ribozomálních genů.
Nově vznikající pre-rRNA v jadérku lidské HeLa buňky byla lokalizována kombinací elektronové a
fluorescenční mikroskopie, přičemž rRNA je jediným typem RNA syntetizovaným v jadérku. K prokázání rRNA
byl použit bromouridin trifosfát (BrUTP), který je začleněn do RNA ve formě bromouridinu. Ke zviditelnění
RNA molekul byla použita imunodetekce bromouridinu protilátkami konjugovanými s koloidním zlatem
(černé tečky v A, B, resp. světlé oblasti v převráceném kontrastu v C, D) či fluorescenční značkou
(červené oblasti v C, D a insert v A). Při znalosti zobrazovací funkce mikroskopu (insert v D) lze
rozlišení fluorescenční mikroskopie zvýšit matematicky dekonvolucí (D, srovnej s C).
Avšak pouze elektronová mikroskopie je v tomto případě schopna správně lokalizovat transkripci
ribozomálních genů jednoznačně do denzních fibrilárních komponent jadérka (d). Dekonvoluční přístup (D)
vytváří nesprávný výsledek. Měřítka: 0,5 µm (A), 0,2 µm (B), 1 µm (insert v D). (Malínský et al.,
J Struct Biol. 2002;140:227-31 a Koberna et al., J Cell Biol. 2002;157:743-8)
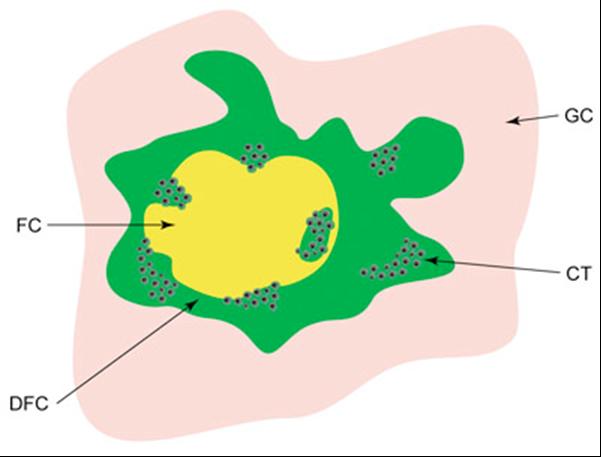 Model uspořádání transkripčně aktivních ribozomálních genů.
Model uspořádání transkripčně aktivních ribozomálních genů.
Schematické znázornění lokalizace nově vznikající pre-rRNA v jadérku savčích buněk. Jednotlivé
kompartmenty jadérka jsou barevně odlišeny. Shluky černých teček (CT) představují jednotlivé
aktivní geny. FC - fibrilární cetrum, DFC - denzní fibrilární komponent, GC - granulární komponent.
(Raška et al., Biol Cell. 2004;96:579-94)
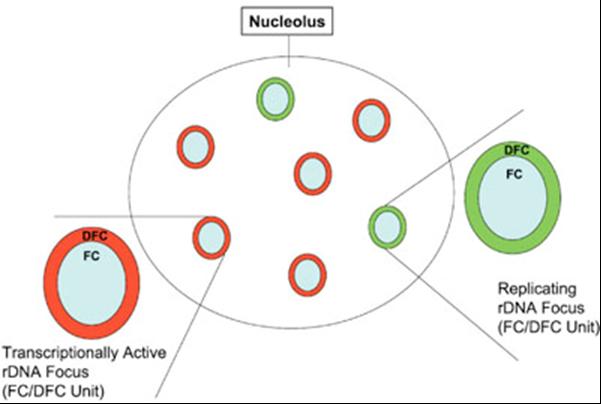 Schéma funkčního zapínání a vypínání ribozomálních genů.
Schéma funkčního zapínání a vypínání ribozomálních genů.
Transkripčně aktivní ribozomální geny jsou soustředěny do denzních fibrilárních komponent (DFC) jadérka.
Během S-fáze buněčného cyklu dochází k nastartování jejich replikace vždy tak, že všechny geny soustředěné
kolem jednoho fibrilárního centra (FC) jsou replikovány najednou. Celá oblast kolem jednoho FC je tak zapojena
buď pro replikaci (zeleně), nebo přepis (červeně) ribozomálních genů. (Pliss et al., J Cell Biochem.
2005; 94:554-65)
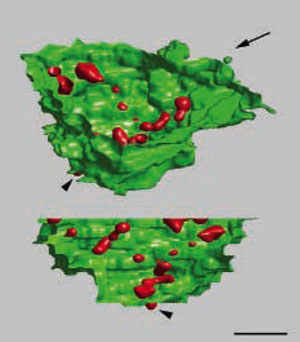 3D lokalizace jadérkové transkripce.
3D lokalizace jadérkové transkripce.
Rekonstrukcí 26 sériových ultratenkých řezů byl sestaven trojrozměrný obraz transkripční aktivity ribosomálních
genů (červeně) uvnitř jadérka (barvička YOYO-1, zeleně) HeLa buňky. Prezentovány jsou dva pohledy, směr spodního
pohledu je indikován šipkou. Měřítko, 1µm. (Koberna et al., J Cell Biol. 2002;157:743-8)
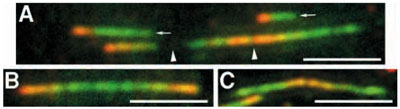 Rychlost replikační vidličky DNA.
Rychlost replikační vidličky DNA.
V živých buňkách byla deriváty deoxythymidinu (chloro- a iododeoxyuridinem) značena nově vznikající DNA.
Po lýze buňky a roztažení DNA bylo možno identifikovat nově syntezované úseky DNA a změřit rychlost replikační
vidličky. Měřítka, 5µm. (Malinský et al., J Cell Sci. 2001; 114:747-50)
 Elektronmikroskopická lokalizace DNA replikace.
Elektronmikroskopická lokalizace DNA replikace.
Nově vznikající DNA buněk v pozdní S-fázi byla naznačena biotinyl-dUTP, modifikovaným deoxythymidin-trifosfátem.
Ten byl následně lokalizován stříbřením imunokomplexu obsahujícím nanočástice koloidního zlata
(černé kuličky představující precipitáty stříbra). Zřetelně patrné jsou jak izolované shluky černých kuliček
(replikační ohniska), tak větší oblasti představující shluky replikačních ohnisek (insert), přičemž jednotlivé
ohniska jsou označena šipkou. Měřítko, 1µm. (Koberna et al., J Cell Biochem. 2005; 94:126-38)
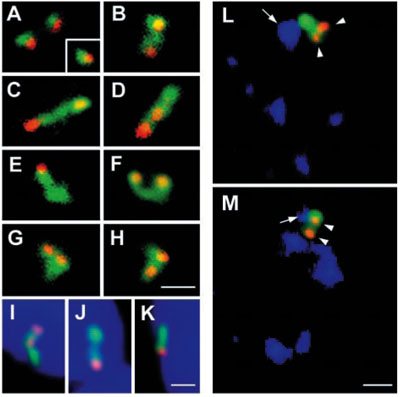 Mapování přepisu specifických genů.
Mapování přepisu specifických genů.
Prostorové mapování genu (červeně) a transkribované RNA (zeleně) na příkladu DNA EBV (viru Epstein-Barrové).
Ve většině případů (B-H) se transkribovaná RNA postupně vzdaluje od genu, pouze v malé části buněk je signál
patrný pouze v místě transkripce (A). Toto putování pre-mRNA nemá žádný vztah k jaderné obálce (I-K, modře je
značeno jádro barvičkou DAPI) a je nasměrováno k jaderným skvrnám obsahující sestřihový faktor SC35
(modře v L, M). Měřítka, 1,5µm. (Melčák et al., Mol Biol Cell. 2000;11:497-510)
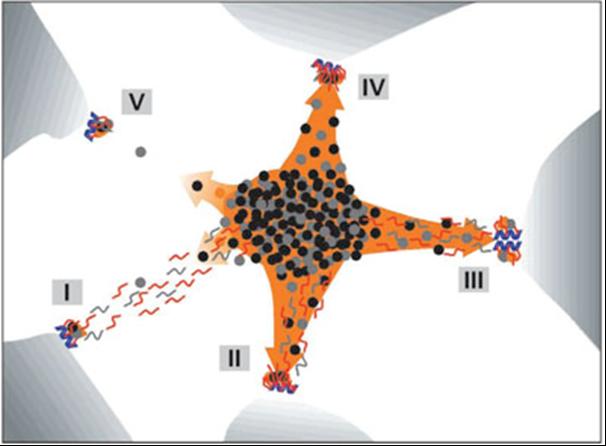 Strukturně funkční dynamické vztahy mezi přepisem genů a sestřihem.
Strukturně funkční dynamické vztahy mezi přepisem genů a sestřihem.
Obraz mapování genů (modré šroubovice), RNA (červené a šedé úseky) a sestřihových faktorů (kolečka) se liší
podle úrovně transkripční aktivity a rychlosti sestřihu. Při mírné úrovni transkripce a převážně posttranskripčním
sestřihu (I) transkripty putují do jaderných skvrn (speckles, oranžová oblast), při převaze kotranskripčního
sestřihu se putující transkripty stávají součástí jaderné skvrny (II). Stejně je tomu v případě silné transkripční
aktivity s oběma typy sestřihu (III). Ve všech třech případech lze pozorovat přesun RNA od genu k jaderným skvrnám.
Pokud dochází k sestřihu pouze kotranskripčně (IV), je RNA soustředěna pouze v místě transkripce. S
jadernou skvrnou jsou pak asociovány jak RNA, tak samotný gen. Tato asociace se vytrácí při nízké úrovni
transkripce (V). (Melčák et al., Mol Biol Cell. 2000;11:497-510)
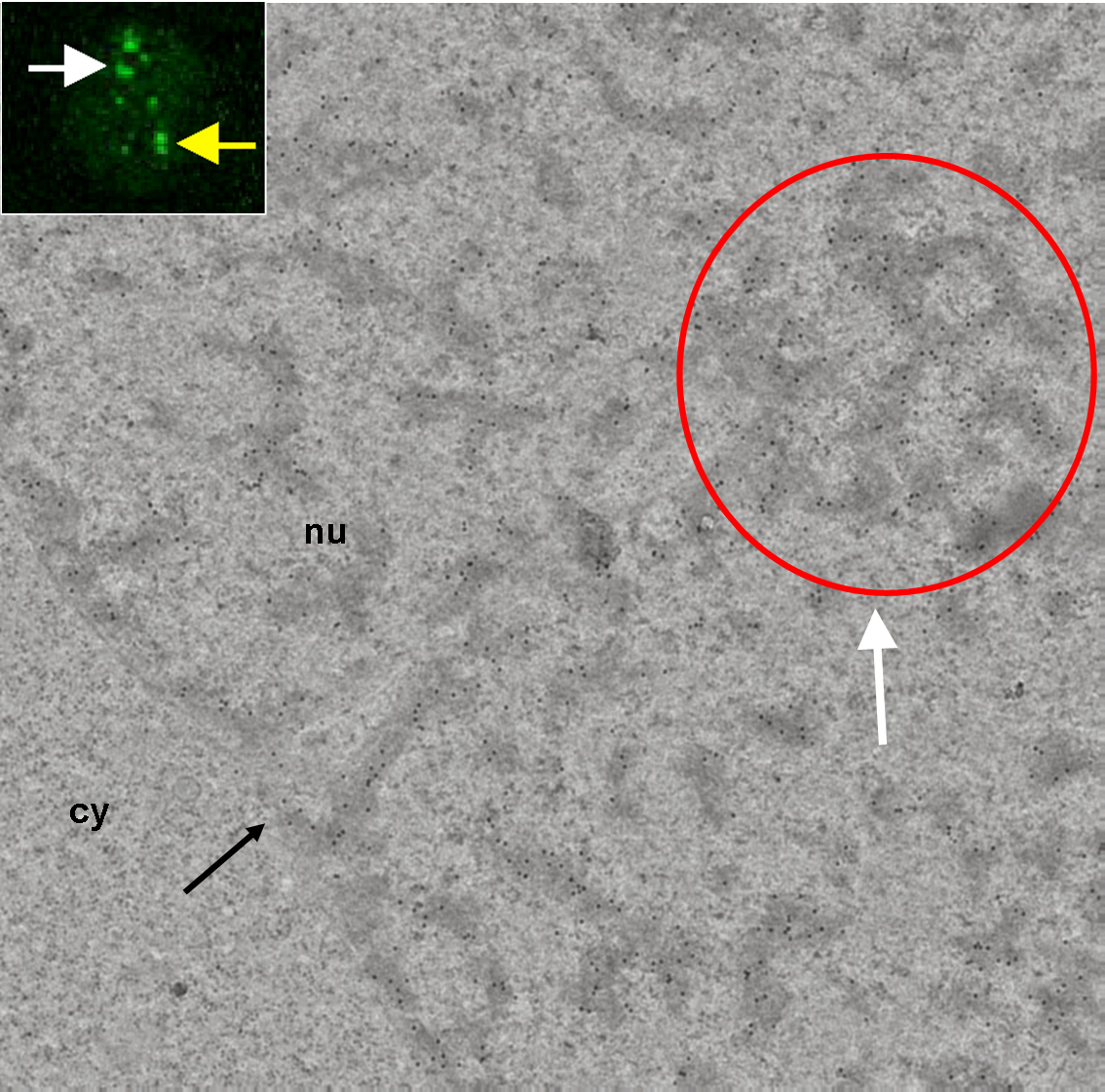
Korelační světelná a elektronová mikroskopie Polycomb tělíska. Fluorescenční obraz Polycomb tělísek vizualizovaných pomocí fůzního proteinu BMI1-GFP. Dvě Polycomb tělíska jsou označena bílou a
žlutou šipkou. Na elektronogramu fluorescenčnímu Polycomb tělísku, ve výřezu označeném bílou šipkou, odpovídá jadérná doména (resp. její řez) s nahromaděním heterochromatinových struktur
(v červeném kruhu). Elektrondenzní heterochromatinové struktury jsou specificky imunoznačeny proti proteinu BMI1. Mimo kondenzovaný chromatin jsme žádné imunoznačení nepozorovali. Co se týče
značení heterochromatinových struktur, jeho denzita je uvnitř a vně „Polycomb tělíska“, vztaženo na plochu heterochromatinu, porovnatelná.
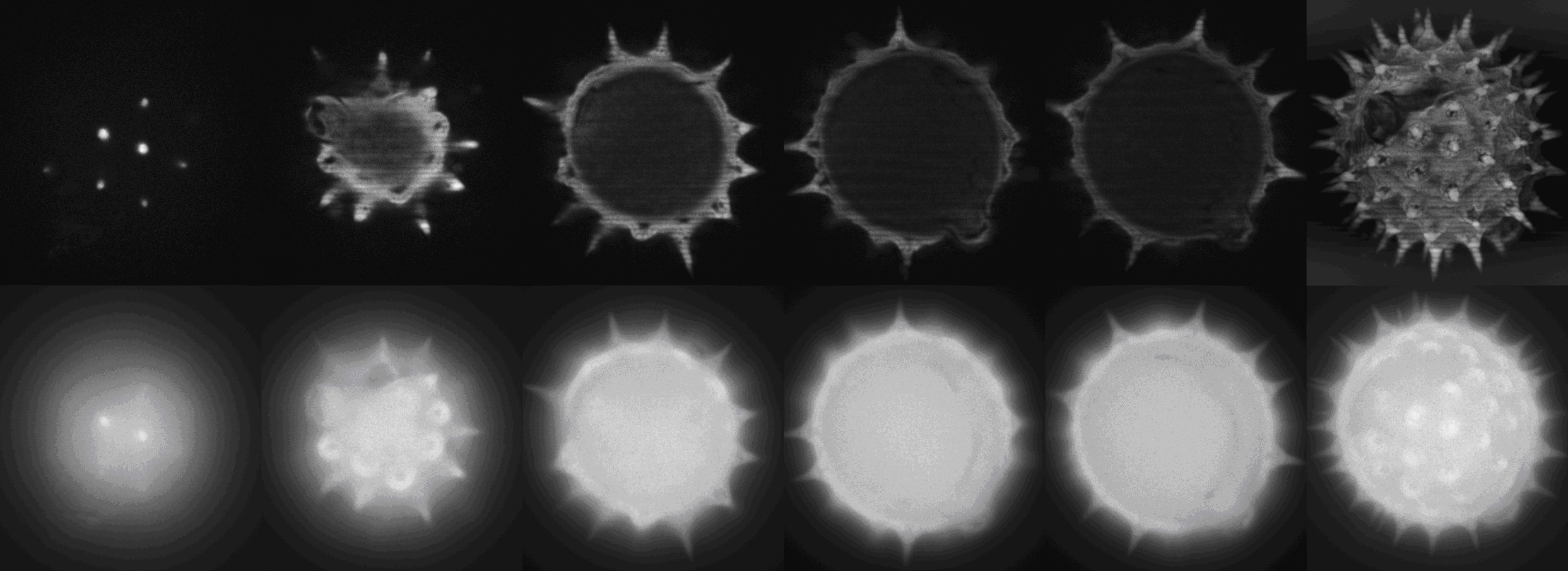 Demonstrace metody optických řezů při snímkování zrna březového pylu.
Horní řada:
Demonstrace metody optických řezů při snímkování zrna březového pylu.
Horní řada: Obrazy získané
strukturovaným osvětlením vytvořeným světelným modulátorem v PAM systému vyvinutém na ÚBBP.
Dolní řada: Obrazy získané konvenční světelnou
mikroskopií.
Panely zcela vpravo srovnávají maximální projekce
získané oběma typy osvětlení.
Hlavní výhodou použití prostorových světelných
modulátorů je výrazné zvýšení rychlosti akvizice obrazů, která je vhodná pro
pozorování živých buněk a vede ke snížení vybělování a fototoxicity.

Superrozlišení docílené PALM/STORM přístupem.
Vlevo: Konvenční fluorescenční snímek A431 buňky syntetizující erbB3-mCitrinové molekuly.
Měřítko: 5 um.
Uprostřed: Snímek v superrozlišení z oblasti vyznačené kružnicí v obr. (a).
Měřítko: 5 um.
Vpravo: Zvětšená oblast vymezená kružnicí v obr. (b). Jednotlivé molekuly
jsou lokalizovány s přesností až 20 nm. Měřítko: 1 um.
 Elektronmikroskopické snímky silika-silikonových nanokapslí připravených metodou
templátové polymerace
Elektronmikroskopické snímky silika-silikonových nanokapslí připravených metodou
templátové polymerace (Kepczynski a spol., 2009, 2010).
Levý panel: zobrazení v konvenční elektronové mikroskopii metodou tzv. negativního kontrastování.
Pravý panel: zobrazení v nativní kryoelektronové mikroskopii.
Snímky ve stejně řadě odpovídají identickým vzorkům. Na první pohled je zřejmé, že obraz v konvenční mikroskopii neodpovídá skutečnému stavu. Měřítko: 100 nm (obrázek vpravo dole má stejné zvětšení jako obrázek vpravo nahoře).
 Elektronová tomografie tenkých nanokompozitních vrstev typu cín/plazmový polymer
Elektronová tomografie tenkých nanokompozitních vrstev typu cín/plazmový polymer (Matoušek a spol., 2009).
Vlevo: Standardní projekce naprášené vrstvy získaná transmisním elektronovým mikroskopem Tecnai Sphera 20 při urychlovacím napětí 200 kV, se zvětšením 25000x. Měřítko: 50 nm.
Vpravo: Tři vzájemně řezy tomografickou rekonstrukcí vrstvy vypočítané ze souboru 69 projekcí naklápěných v rozsahu ±70°. Řezy jsou zobrazeny s obráceným kontrastem.
Dole: Trojrozměrná vizualizace zrekonstruované vrstvy.
Tenká vrstva byla připravena magnetronovým naprašováním z cínového terčíku ve smíšené atmosféře par argonu a n-hexanu.

